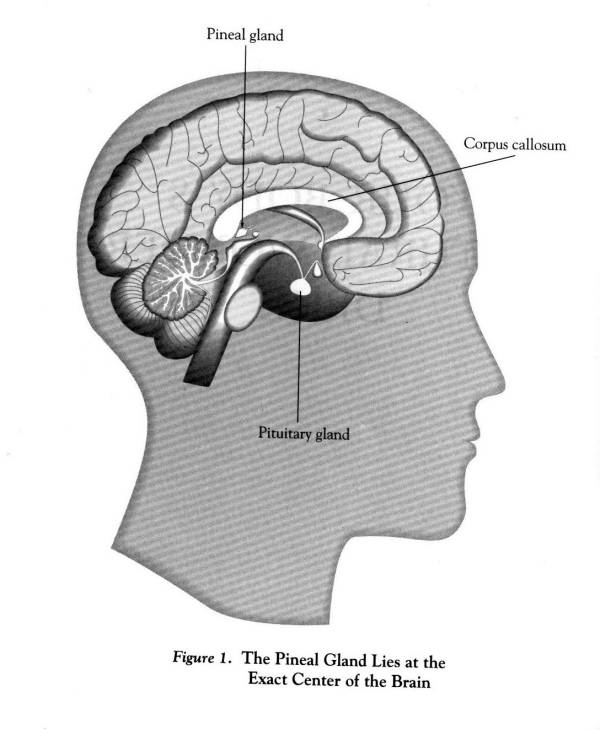
STRUCTURE OF THE PINEAL GLAND
At first sight, the structure of the mammalian pineal
is not very exciting. It lies at the exact centre of the brain behind the
eyes as shown in Fig.1 above and consists of only two major cell types,
pinealocytes and immature astrocytes. Occasionally, the pinealocytes are
arranged in follicles surrounding narrow or wide spaces. The gland is richly
innervated by postganglionic sympathetic nerve fibres, most of which are
found in the perivascular spaces of capillaries. In addition, pinealopetal
fibres of central origin are present (Vollrath 1981). Observed under the
light microscope the pineal specific cells, the pinealocytes, lack prominent
and differential staining properties. Special staining reagents such as
silver impregnation are necessary to demonstrate their complete outlines.
Then it becomes apparent that pinealocytes are nerve cell like, consisting
of a perikaryon and an unknown number of cytoplasmic processes. The large,
pale nucleus with its prominent nucleolus is also reminiscent of a large
ganglion cell. As cytoplasmic basophilia is virtually non existent,
there is no satisfactory explanation for the high metabolic activity of
pinealocyte nuclei.
At the ultra structural level it is quite difficult to relate structure
to function. In most mammalian species investigated the pinealocytes contain
few granules that can be regarded as morphological correlated of
secretory products, and the circadian behaviour of the few dense core vesicles
present does not support the assumption that they contain melatonin. According
to Julliard & Collin (1980) dense core vesicles act as storage sites
for serotonin. Pinealocyte perikarya house a prominent Golgi apparatus
and relatively small amounts of smooth and rough endoplasmic reticulum.
In some species cisternea of the endoplasmic reticulum contain flocculent
material which may represent a form of secretory substances. Highly
pleomorphic mitochondria tend to form clusters reminiscent of the ellipsoids
of inner segments of photoreceptor cells. That the pinealocytes are responsible
for the conversion of serotonin to melatonin has been demonstrated histochemically
by showing that it is these cells that contain serotonin (Bertler et al
1963) and melatonin (Freund et al 1977) and not the astrocyte-like interstitial
cells.
It is generally accepted that mammalian pinealocytes are phylogenetically
derived from pineal photoreceptor cells. In lower vertebrates pineal photoreceptors
are similar to retinal cones and show outer and inner segments as well
as synapses with afferent pinealofugal nerve fibres. During phylogenesis
the outer segments regress as do the pinealofugal nerve fibres, but the
synaptic ribbons of the afferent synapses persist. In view of the phylogenetic
regression and the current concept that it is the function of mammalian
pinealocytes to synthesize melatonin and to release it into the systemic
circulation, both the shape of the pinealocytes and the architecture of
the gland are surprising.
Human pinealocytes are equipped with long cytoplasmic processes. Comparable
processes are present in all mammalian species investigated ultra structurally.
When we compare these process bearing cells with the pineal photoreceptor
cells of lower vertebrates it is difficult to envisage that mammalian pinealocytes
represent regressed photoreceptor cells. Instead it appears that mammalian
pinealocytes are highly differentiated cells similar to nerve cells, the
process of which receive messages and pass on signals. A puzzling feature
is that, although the possible morphological correlates of pineal secretory
products are particularly prominent in terminal swellings of pinealocyte
processes in many mammalian species, only a few terminals are close to
blood vessels. Instead, the perivascular spaces are filled with large bundles
of postganglionic sympathetic nerve fibres. In fact, according to
quantitative studies in the rat, 91.1% of the nerve fibres have a perivascular
location, the remainder lying between pinealocytes (A.Meyer & L. Vollrath
1985). It is enigmatic that sympathetic nerve fibres predominate in the
perivascular spaces since the nervi conari reach the pineal gland independently
of blood vessels, as separate nerves.
INNERVATION OF PINEALOCYTES
Sympathetic innervation
This type of innervation has been clearly defined both morphologically and functionally. The fibres originate in the superior cervical ganglia (SCG) of the sympathetic trunk, continue in the internal carotid nerve and enter the pineal gland as nervi conari (Zigmaond et al 1981, Bowers et al 1984). Their importance for the regulation fo melatonin synthesis has been demonstrated by biochemical studies after sympathectomy or electrical stimulation of the SCG, the latter leading to an approximately 50 fold increase of serotonin N-acetyltransferase (NAT) activity (Bowers & Zigmond 1980,1982). A study of rat pinealocytes after electrical stimulation of the SCG by Beuss in 1985 reviewed that some pinealocytes did not appear to be influenced by SCG stimulation, a second group responded with enhanced electrical activity and in a third group electrical activity was depressed. In view of the continuing controversy about whether NAT of hydroxyindole O-methyltransferase (HIOMT) is the rate limiting enzyme for melatonin synthesis, and the lack of a clear-cut day / night rhythm of HIOMT, in contrast to NAT, it is relevant to recall that as early as 1972 it was reported that preganglionic electrical stimulation of sympathetic nerves resulted in an increase of pineal NAT activity but a decrease of HIOMT activity.
Central innervation
Nerve fibres reach the pineal gland via habenular and posterior commissures
and now with modern neurobiological techniques available, it becomes
apparent that these fibres are of functional importance. Lesion and horseradish-peroxidase
studies have revealed that central pinealopetal nerves fibres originate
in diverse brain regions including the habenular, paraventricular and suprachiasmatic
nuclei as well as the preoptic area, amygdala, olfactory centres, lateral
geniculate bodies and the sites of origin of the stria medullaris. The
central fibres contain a variety of peptides such as oxytocin, vasopressin,
luteinizing hormone-releasing hormone, vasoactive intestinal polypeptide.
The fibres are unevenly distributed in the pineal gland, some lying in
the periphery and others in the centre.
FUNCTION OF THE PINEAL GLAND
The mammalian pineal gland evolved from a well differentiated photoreceptive
organ in lower vertebrates, a functional third eye ![]() .
In human, pineal gland is a small pea size structure situated in the middle
of the brain. The mean weight of the pineal in a woman is 173 mg which
does not differ from the mean weight of 172 mg for a man
.
In human, pineal gland is a small pea size structure situated in the middle
of the brain. The mean weight of the pineal in a woman is 173 mg which
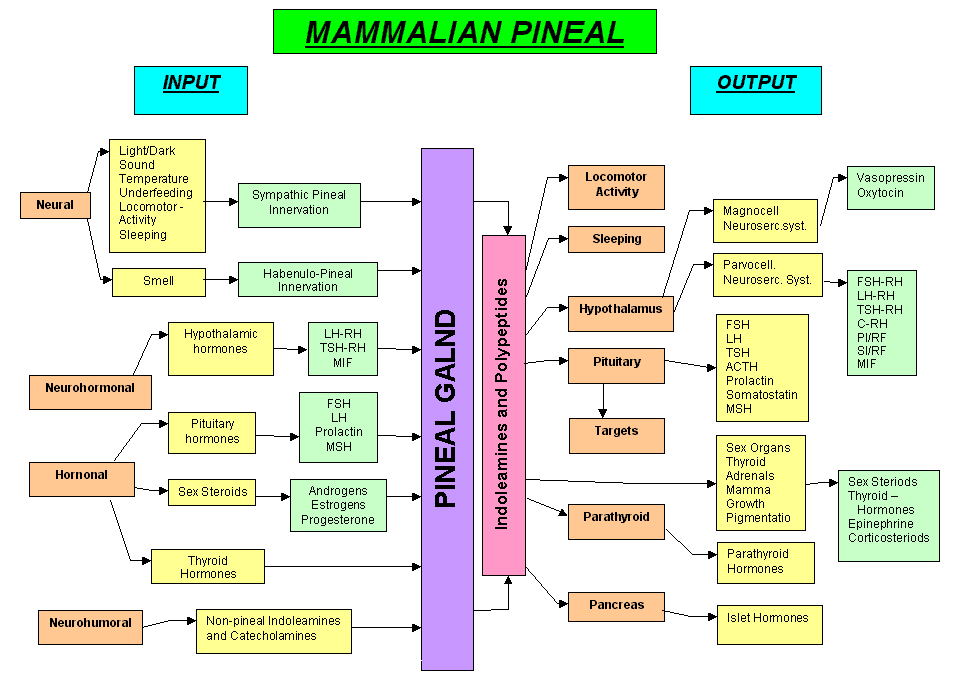
does not differ from the mean weight of 172 mg for a man ![]() . The function of the pineal gland is shown in Fig. 2. Several conditions
and compounds arranged in four categories have been listed which are known
to influence pineal function. Conditions of stress could affect the pineal
either by way of the pineal sympathetic innervation of via the neurohumoral
route. On the right or output side, indoleamines and polypeptides are mentioned.
These are substances include melatonin which are known to be produced and
generally secreted by the pineal.
. The function of the pineal gland is shown in Fig. 2. Several conditions
and compounds arranged in four categories have been listed which are known
to influence pineal function. Conditions of stress could affect the pineal
either by way of the pineal sympathetic innervation of via the neurohumoral
route. On the right or output side, indoleamines and polypeptides are mentioned.
These are substances include melatonin which are known to be produced and
generally secreted by the pineal.
The pineal gland does not have the capacity to respond directly to light. Rather, light controls it through a system which includes the lateral eyes, central and peripheral neural structures and neurochemical transduction mechanisms within the gland. All vertebrates appear to synthesize melatonin rhythmically on a 24h basis. Click here to see the amount of melatonin secretion by the human pineal gland during a 24 hour cycle.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
The effect of light on the melatonin rhythm generating system are properly thought of as effects on the SCN. Although the SCN can function in a cyclic manner autonomously, environmental lighting has strong effects on the changes in pineal N-acetyltransferase ( the enzyme that converts serotonin to melatonin ) activity and melatonin production. The SCN clock is actually composed of two dependent clocks and the degree to which their pineal stimulatory periods overlap is determined by the amount of light. Long nights appear to allow the clocks to drift apart, so that the pineal gland will be stimulated for a longer period than in animal kept in short nights. The effect of exposure to light is also to determinate the neural stimulation of the pineal gland. This results in the decrease in the enzyme N-acetyltransferase and therefore melatonin production. For the biological synthesis of melatonin
The amount of melatonin produced is directly linked to the sleep pattern
in mammals including human. The raw material used to made melatonin is
the amino acid tryptophan (chemical name). The tryptophan we consume
during the day is converted into serotonin (chemical name), a brain chemical
involved with mood. Serotonin in turn is converted into melatonin. The
action of melatonin and serotonin have profound effects on homeostasis,
immune surveillance and the maintenance of connective tissue, constructural
and muscular components.
Other interesting biological properties of melatonin are listed as follow;
The degenerative processes associated with aging is a biologically complex and multifaceted phenomena. A current, but putative theory on aging associates the gradual accumuation of oxidative stress in neural tissue to accelerated neurodegenerative changes and age-related diseases. Oxidative stress is defined as the cellular damage caused by oxygen free radicals. The two types of oxygen free radicals, the superoxide anion and the hydroxyl radical are naturally produced by product of aerobic metabolism, the latter bding more biologically toxic. The reactive nature of free radicals stems from their unpaired valence electron that mediates oxidative toxicity, damaging nucleic acid, membrane lipids, proteins, and carbohydrates. Approximately 5% of celular oxygen is not used in the production of ATP but is reduced to reactive free radicals. An estimated 1011 free radicals/cell/day formed, inducing perhaps up to 105 oxidized DNA residues formed/cell/day. Furthermore, it is suggested that the progression of brain cell aging occurs when the balance between oxidative stress and antioxidative defense tips towards increased free radical production. Neural activity,especially the release of the accumulation of oxidative stress leads to the morphological and physiological destruction of neurons. A gradual deterioration of neurological tissue occurs represented by functional loss, such as slowed reactions, diminished memory, or tremour. This progressive degeneration is the priced the ody pays for utilixing oxygen. Increased free radical generation may also results from the additive exposure to toxins, UV light and stress or from the decrease in the bodies defense sysems to reduce oxidative stress, namely free radical scavengers, antioxidative enzymes, or meal chelating agents.
Melatonin's Protecting Effect
Melatonin's protecting role in aging lies in its non-receptro mediated
interactions as a potent oxygen radical scavenger. Studies have shown that
endogeneous levels of melatonin have oxidative protective effects, with
a greater reduction in DNA damage at night, reflecting melatonin's phasic
secretion pattern. It scavenges both the superoxide anion and the hydroxyl
redical, protecting nuclear DNA, proteins and membrane lipids against free
radical damage, as well as stimulating the activity fo glutathione peroxidase,
putatively the most important antioxidant in the brain. The indole's lipo
and hydrophilicity are properties unique to antioxiidants and allow for
rapid diffusion and accessibility to all subcellular components as well
as freely crossing the blood-brain barrier. Other antioxiiidants are confined
to particular cellular compartments such as lipid cell membranes for vitamin
E and the cytosol for vitiamin C.
The pineal gland as an integral constituent of
the neuroendocrine system seems to play an important role in modulating
the immune response via circadian release of its main neurohormone melatonin
and/or some other substances. There is a substantial body of evidence suggesting
an antimitotic action of melatonin in mammalian cells in vitro![]() .
.
Possible mechanisms of interacting between the immune
and the pineal gland hormones seems to come full circle with the pathophysiology
of the immune disorders. Neuopeptides and neurotransmitters such as endorphins,
enkephalins, vasoactive intestinal peptide and the pineal hormones all
have significant influence on the immune system, and have been shown to
modulate antibody production, natural killer cell activity and response
to mitogen. The products of the immune system, on the other hand, has substantial
influence on the pineal gland and neuroendocrine system.
Similar antineoplastic effect of the pineal gland and its hormone melatonin
was observed in fibrosarcoma, in a transplantable form of leukaemia and
various form of carcinoma. The enhancement of transplantable tumour growth
in pinealectomised animal has been reported long time ago.
Modern investigations have revealed that about 50% of pineal tumours in
humans are germinomas. Pineal tumours are most common in the Japanese population
and occur four to five times more often in males than in females![]() .
In the United States, as many as 40% of tumours in the region of the pineal
gland are a mixed historical type
.
In the United States, as many as 40% of tumours in the region of the pineal
gland are a mixed historical type![]() .
However, studies on the link between the pineal gland and
tumour development did not always yield consistent results, though many
of the reports pointed to oncostatic action of the pineal. There have also
been several papers reporting that melatonin has no or even stimulatory
effects on the growth of some tumours Differences in the results obtained
may depend on a number of reasons. In fact, precise comparison of the studies
on relationship between the pineal neurohormones and neoplastic growth
is very difficult due to the diversity of the experimental approaches i.e.
various tumour models used, different methods of measurement of tumour
growth (neoplastic cells proliferation, tumour weight, tumour volume),
differences in mode and timing of melatonin administration and various
photoperiodic environment. In general however, most results have pointed
toward an inhibitory effect of melatonin on tumourigenesis
.
However, studies on the link between the pineal gland and
tumour development did not always yield consistent results, though many
of the reports pointed to oncostatic action of the pineal. There have also
been several papers reporting that melatonin has no or even stimulatory
effects on the growth of some tumours Differences in the results obtained
may depend on a number of reasons. In fact, precise comparison of the studies
on relationship between the pineal neurohormones and neoplastic growth
is very difficult due to the diversity of the experimental approaches i.e.
various tumour models used, different methods of measurement of tumour
growth (neoplastic cells proliferation, tumour weight, tumour volume),
differences in mode and timing of melatonin administration and various
photoperiodic environment. In general however, most results have pointed
toward an inhibitory effect of melatonin on tumourigenesis![]() .
.